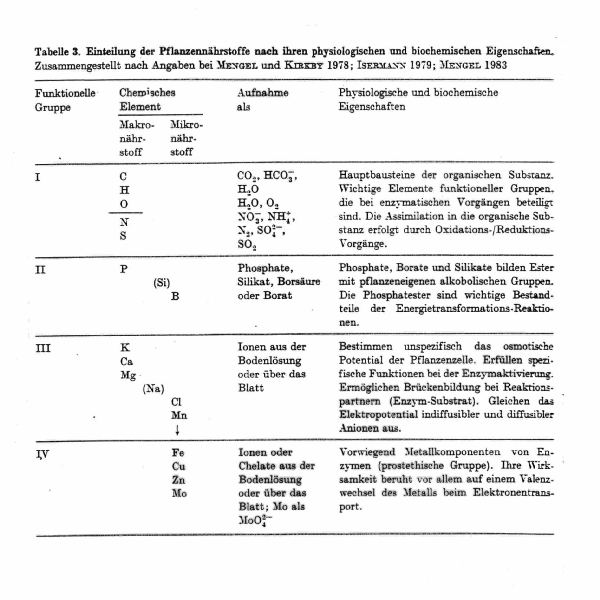
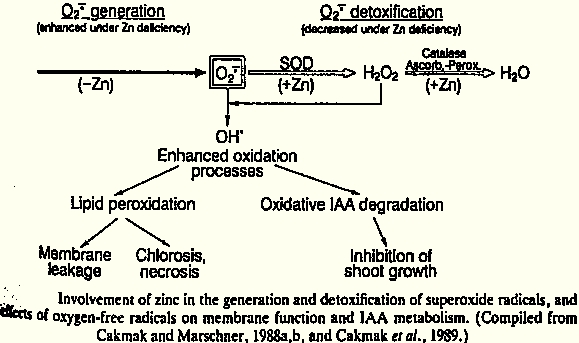
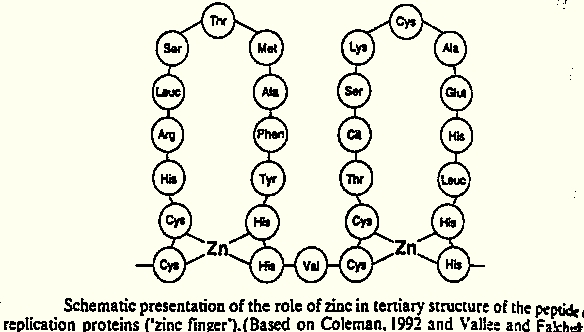
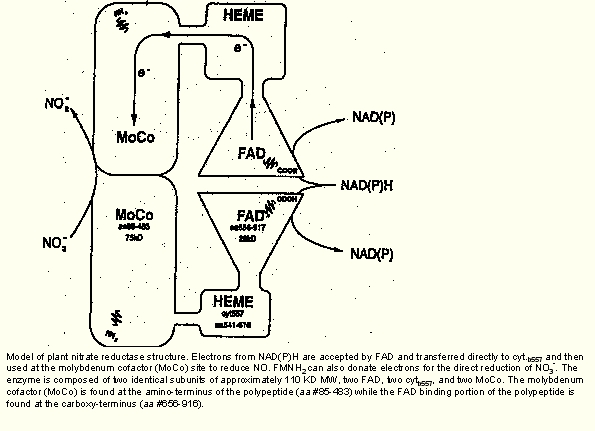
This group has functions in regulating osmotic and ionic balances. Beyond that, it
has specific
roles in enzyme conformation and catalysis.
Example: Calcium
Excerpt from: Calcium as a "second messenger." Source: http://www.biologie.fu-
berlin.de/lampart/Seminar2000/CALCIUM.HTM
Calcium is required, among other things, for maintaining the structure and function
of all cell
membranes. Within the cell, calcium as a divalent cation (similar to Mg²⁺) can form salts
with acidic
membrane components (e.g., protopectin in middle lamellae, the walls of root hairs and pollen
tubes, or alginic acid in algal cell walls) and thus serve as an essential building block.
For example, Ca²⁺ deficiency inhibits pollen germination and pollen tube
growth. Furthermore, this
deficient state leads to damage of meristems, especially root meristems. Monocots require less
Ca²⁺ for optimal growth than dicots.
Ca²⁺ also serves as a (relatively non-specific) cofactor for a number
of enzymes and acts in
balancing the effects of other cations.
For these aforementioned functions of calcium, a much lower concentration would generally
suffice
than is normally found in plants. The excess Ca²⁺ is fixed within the cell as a poorly soluble
salt in
the form of, for example, phytate, oxalate, carbonate, or (more rarely) sulfate or phosphate.
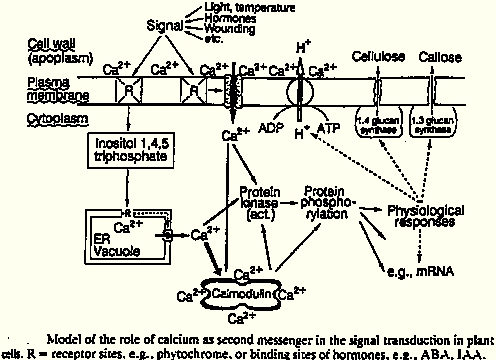
Free cytosolic Ca²⁺ ([Ca²⁺]c) is a convergence point for many
different signaling pathways. Each
signal has its own Ca²⁺ "fingerprint," a spatially unique structure comprising
specific combinations
of several hundred proteins that then act together. Certain combinations of ion flux changes and
gene expression support the subsequent physiological response. In Ca²⁺ changes, a distinction
is
made between degenerative processes, like senescence, and adaptive responses, which are
crucial for signaling pathways.
Under the influence of various external factors, e.g., touch, cold, wind, elicitors,
hyperosmotic
shock, there is an increase in the cytosolic Ca²⁺ concentration from < 2µM (at rest)
to 1-10 µM in
the excited state, partly involving the Ca²⁺/calmodulin system. The influx of Ca²⁺
upon stimulation
occurs from the vacuole, the ER, and the apoplast, primarily likely via voltage-dependent ion
channels. The efflux of Ca²⁺ from the cytosol and thus the restoration of the resting state
is
accomplished by Ca²⁺- ATPases in the plasma membrane and ER, a Ca²⁺/nH⁺
antiporter in the
tonoplast, and a Ca²⁺ uniporter in the chloroplast membrane.
Signaling Pathways
Signaling pathways consist of the perception of signals of a physical or chemical
(e.g., hormones)
nature, their intracellular transduction, and the response. The receptor should traverse the plasma
membrane to receive the signal from outside and relay it to the cytoplasm.
A guanine nucleotide-binding protein ("G-protein") can activate a further
signal transduction chain,
thereby converting extracellular signals into intracellular ones. An example of this is the
phosphoinositide cascade. The messengers involved here are generated from phosphatidylinositol-
4,5-bisphosphate (PIP₂), a phospholipid of the plasma membrane. An activated phospholipase
C
(PLC) in the cytosol hydrolyzes PIP₂ into two "second messengers": inositol-1,4,5- trisphosphate
(IP₃) and diacylglycerol (DAG). PLC activity is significantly increased by a rise in the cytoplasmic
Ca²⁺ concentration. This increase, in turn, is achieved by the opening of channels for Ca²⁺
in
intracellular stores (e.g., ER) under the influence of IP₃.
Signal Transmission in the Whole Plant
Tobacco seedlings, under continuously high temperatures, induce short-lived [Ca²⁺]c
transients
(currents) lasting 20-25 minutes. New heat shock [Ca²⁺]c transients can only be induced again
after
a recovery period of 8 hours at ambient temperature. During this recovery period, normal [Ca²⁺]c
transients can still be induced by cold and wind signals. Heat, cold, and wind use different
transduction pathways or mobilize spatially distinct pools of [Ca²⁺]c to produce a signal-specific
[Ca²⁺]c fingerprint.
The kinetics of [Ca²⁺]c transients are unique for each signal: the lag
phase before the current, the
subsequent increase in Ca²⁺ concentration, the peak of the current, and the decline back
to the
resting state.
Possible targets of these currents could be plasma anion channels and a K⁺
release from the
vacuole.
Literature
Bowler, C., Neuhaus, G., Yamagata, H., Chua, N. (1994) Cyclic GMP and Calcium Mediate
Phytochrome Phototransduction. Cell, 77: 73-81
Lüttge, U., Kluge, M., Bauer, G. (1994) Botanik. VCH. 2nd Edition, pp. 505-507
Neuhaus, G., Bowler, C., Hiratsuka, K., Yamagata, H., Chua, N. (1997) Phytochrome-regulated
repression of gene expression requires calcium and cGMP. EMBO J, 16, No. 10, pp. 2554-2564
Sitte, P., Ziegler, H., Ehrendorfer, F., Bresinsky, A. (1998) Strasburger Lehrbuch
der Botanik.
Gustav Fischer, 34th Edition, pp. 326, 366-368
Trewavas, A., Malho, R. (1998) Ca²
⁺ signalling in plant cells: the big
network! Curr. Opin. Plant
Biol., 1:428-
433